 区块链神吐槽
区块链神吐槽常有人感叹人类食谱之广泛,简直能把什么东西都弄上餐桌,从某些角度看,确实如此,不过这里有几件容易混淆的事情,首先,人类食谱之广泛,主要归功于人类文化的巨大多样性,群体间的饮食习俗差异,以及个体间的口味嗜好差异,假如分解到单个群体或个人,其广度就远不如一本《食材大全》所显示的那么值得惊叹了。
其次,假如我们随便挑几个食俗不像因纽特人那么极端的群体,用赫芬达尔-赫希曼指数(Herfindahl-Hirschman Index)——这是经济学家度量供方离散度的标准方法——来测量食物来源离散度,那么人类得分确实不低,毕竟我们是杂食动物,可是,假如我们把衡量标准换成『有能力消化因而有可能吃多少种食物并从中获取营养』的话,那么得分最高的脊椎动物远不是人类,而是——你或许会吃惊——食草动物(注:除非特别说明,本文所谈论的动物仅限于脊椎动物)。
食草动物也吃肉
因为凡食肉动物和杂食动物吃的东西,食草动物几乎也都能吃,鹿经常被观察到在啃动物尸体,甚至同类的内脏,牛在吃草叶时也常有开点小荤的机会:草丛里的蜗牛,树上掉下来的雏鸟或鸟蛋,死老鼠……河马上岸吃草时甚至偶尔会主动猎杀动物,畜牧业者也早就懂得往牛羊饲料里添加屠宰下脚料。

食草动物不会放过可以吃肉的机会
反过来却不行,食草动物消化纤维素和对付植物毒素的能力太强大了,以至很多被它们当作主食的植物其他动物都吃不了,而食草动物很少吃其主食之外的东西,特别是肉食,并不是因为消化吸收上存在任何障碍,而是一种策略选择:基于它们在生理和技能上的相对优势,把时间和精力花在寻找、争夺和获取肉食上,几乎总是不划算的。
比如一头鹿,在一天中可用于觅食的那几个小时里,若面临两个选择:要么专心吃树叶(并时刻警惕着随时出没的老虎),要么漫游林中寻找尚未过度腐烂的动物尸体,动物尸体能量密度高,消化成本低,一只野兔或许顶得上啃两天树叶的净收益,可同时,大张旗鼓的搜索尸体,扩大了活动范围,提高了自己的活跃度和曝光率,因而更可能被老虎吃掉,也增加了与食腐动物(比如狼)发生冲突的机会。
为了避免与狼冲突,食草动物才选择吃草
更要命的是,搜索尸体的结果远不如啃树叶那么确定,很可能连续几天一无所获,况且鹿又不像食腐动物那样具备远距离发现尸体所需要的灵敏嗅觉,也不像老虎那样能够大块吞肉,一次吃下一周所需,相比之下,树叶虽能量密度低,消化成本高,但收益十分确定,因其难消化,竞争者也少,而且竞争者都是无威胁的食草动物。

对鹿而言,吃树叶是稳定的能量来源
只有当尸体是沿途偶遇的,并且附近没有危险的竞争者,因而无须承担上述种种风险时,鹿才会去吃,这就好比偷窃,食肉动物是职业小偷,将生计建立在偷窃之上,并为此而发展了高度特化于偷窃的生理机制、行为模式和后天技能,食草动物没有这些优势,但若是有顺手牵羊的便宜机会出现,它们也不会漠然放过。
草饲料与谷饲
所以,尽管食草动物拥有强大的纤维素消化和毒素处理能力,但只要在成本与风险无差异的条件下给它们选择,它们还是会偏爱高能量密度、低消化成本和低毒性的食物,野生条件下,成本风险无差异这个条件只是偶尔会满足,而在人工饲养时,由于这些成本和风险转移给了饲养者,而后者拥有的技术又将它们降至极低水平,因而可以轻松满足。
谷物让牛长得更快
于是我们有了谷饲牛,与草饲相比,谷饲牛长肉快,产肉多,脂肪含量高,容易出雪花,同等产肉量所需土地面积仅为草饲的1/3,这些优点对于谷物充裕而草场相对稀缺(相对于加拿大、澳洲和阿根廷)的美国尤为显著,西欧谷物和草场都稀缺,所以更倾向于往饲料里添加屠宰下脚料,这也是为何疯牛病首先在西欧爆发的缘故。
有些情况下,谷饲不仅有好处,而且不可或缺,比如军马;若是只吃草,马一天至少要花八小时咀嚼草料(这是人工饲喂干草的情况,若自己在草场吃,需十几个小时),每公斤嚼3500-4500下,约需40分钟,而且吃完后三四小时内消化负担极重,其长达20米的小肠在此期间将分泌100多升消化液,随后50多升食糜进入一米多长的盲肠并在那里开始发酵。
喂马吃干草,马一天要花8小时咀嚼
这样,每天能用于行军(牵引或骑乘)和作战的时间就十分有限,最多四五个小时,这还得益于马的睡眠很短,每天不到三小时,外加两三个小时的伏坐休息,所以它们能在夜晚继续进食;但若能将部分草料换作谷物,比如燕麦,每公斤咀嚼次数便降至850次,只需十分钟,替换一半即可省下三小时进食时间,并大幅减轻消化负担。
正是谷饲,让优良役马在轻负荷条件下每天能工作多至8-10小时,从而让一些骑兵部队能以每天50-60公里的速度行军(如果能沿路获得补给的话),勉强超出罗马步兵自带给养的行军速度,至少马不再是行军速度的瓶颈。
最优觅食原理
关于特定动物吃什么,不吃什么,偏爱哪些食物,优先寻找哪些食物,当条件改变时食谱会如何改变,以及有关动物食性的其他种种问题,生物学家发展了一套被称为最优觅食理论(optimal foraging theory, OFT)的成本收益分析方法来寻找解释,该理论考虑的因素主要有:觅食的时间成本,失败的几率和自身的风险承受能力,各种食源的竞争强度和自身的竞争优势,在消化能力和消化成本上的相对优势,因暴露在觅食环境中而被捕食的风险,中毒风险,等等。
食草动物面临被捕食的风险
理解该理论的一个要点是,某种动物花最多时间和努力去寻找,因而事实上也吃得最多的,未必是(且常常不是)它最喜爱的食物,反之,它很少或根本不花精力去寻找某些食物,未必是它消化不了、不爱吃、或没能力获取,而常常是因为,在综合考虑上述因素之后,它“发现”,把时间精力投入在寻觅该食物上并不合算,要么失败风险高的难以承受,要么边际净收益低于将这份时间精力投入于其他食源的收益。
沿着这条思路,不同动物的食性差异,觅食相关的种种行为模式,以及人类饮食习俗的形成与变迁,都将得到更为深入也更系统化的理解。
食草与食肉
这是最鲜明的一组对比,但这对名称本身并未揭示出这一对比的要点所在,关键区别其实并不在于食物来自植物还是动物,而在于对待风险的策略差异:食草动物代表了策略光谱的稳妥保守一端,而像猫科这样的顶级食肉动物则代表了冒险激进一端。
捕猎是高风险活动,专以捕猎为生更是高风险生存策略;捕猎成功率往往很低,而且越是大型猎食者越倾向于大型猎物,而猎物越大,成功率越低,猫科之王老虎的成功率只有5-10%,北极熊10%,狼14%,非洲狮18%,体型苗条的猎豹成功率高的出奇,40-50%,但猎物经常被抢走;对于大型猎食者,连续几天空手而归的情况很平常,他们就像赌场里喜欢博大输赢的赌客,赢上一把够吃上一阵,但经常输个精光。
相比之下,草虽然营养密度低,摄食时间长,消化负担重,但分布广泛,供给充分,收益非常确定,一份付出一份回报,是勤恳吃苦耐劳者可以依靠的生计来源;但具备这些特征的食物未必来自植物,在海滩捡拾贝类,在蚁穴舔食蚂蚁或白蚁,在河流入海口捕捞洄游鱼群,都更像是采集而非捕猎,那些以此为生的动物,在生理特征和行为模式上更靠近食草动物。
比如在食蚁兽身上,你看不到食肉动物的典型特征:大脑发达,认知能力强,活跃好动,好奇心强,爱探索,爱玩耍,反倒有许多食草动物的特征:安静,不好动,重防御,以及高度特化的摄食与消化系统:能快速伸缩的超长舌头,高粘度的唾液,胃内用于碾碎昆虫的搓板状结构(类似鸟类的嗉囊),分泌的胃酸是甲酸而非常见的盐酸;类似的,以洄游鱼群为主食的人类族群,其文化与社会结构的各方面都更像农耕者而非狩猎者。
 食蚁兽更似食草动物,安静、不好动
食蚁兽更似食草动物,安静、不好动
风险策略上的分化,起初可能只是因为所处环境不同,比如在空旷平坦的大草原上捕猎,比在温带森林中困难的多,因为最普遍的捕猎方式是偷偷靠近然后突然袭击,老虎和豹在扑袭之前通常会贴近猎物到十几米甚至几米以内,这一战术需要有足够多的掩蔽物,树丛、土丘、岩石、沟壑,或特别高的草,只有像猎豹这样速度优势极为显著的猎手才会在百米之外就发动进攻,或者像非洲野狗这样的团队捕猎者,能靠合作与耐力长途追逐猎物将其拖垮;总之,在那些捕猎难度过高的生态位(另一个例子是热带雨林的树冠层)中,动物更可能向保守稳妥的食草、食果或食虫发展,反之亦然。
 空旷的大草原上,更难狩猎
空旷的大草原上,更难狩猎
然而一旦策略分化出现并长期持续,进化机制便会将这一差异扩大并固化下来,选择压力作用之下,主吃低营养密度食物的动物,重点发展消化和防御,而食肉动物则重点发展搜索、追踪、潜行、擒杀等捕猎技能,这些特征,相信大家都已耳熟能详,这里我仅以视觉为例略加说明。
同样是两只眼睛,食草动物更注重视野广度,以便全方位探知正在靠近的捕猎者,所以双眼分别朝向头部两侧,视野重叠少,比如牛的视野广度330度,重叠部分仅20-50度,马的视野350度,重叠65度,而食肉动物则高度依赖立体视觉和景深感知,因而两眼向前,形成双眼视觉,视野广度小,但重叠度大,比如猫的视野200度,重叠140度,这一差别,在食肉猛禽和其他鸟类的对比中也可看到。
随着时间推移,选择了不同觅食策略的动物逐渐被自然选择朝着不同方向改造,从生理构造,行为模式,到生存技能,都发生了与其主要食物对应的特化适应;这些改造是全方位和成套出现的,因为生理结构受着基本生化规律和长期积累的进化包袱的严格局限,其设计空间有限,要强化某方面性能,就不得不在其他方面作出牺牲,比如发达的消化系统往往对应着较小的大脑和较低的认知能力。
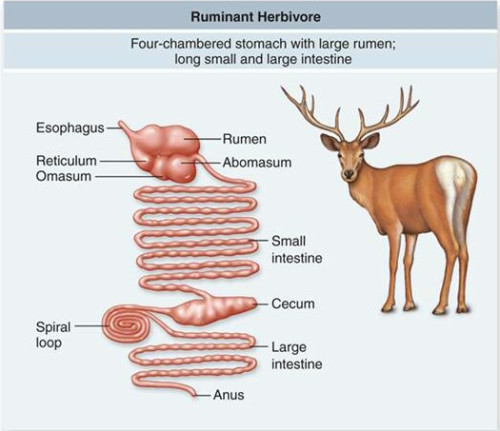 食草动物的消化道很长
食草动物的消化道很长
而且很多生理/行为特性是连锁的,一个改动将引发一系列相应改动,而食性改变往往是触发连串改动的初始启动因素,因而它总是我们认识一种动物生理、习性和行为模式——以及,对于人类,文化与社会结构——的最佳起点;比如在开阔草原吃草的动物都成群出没,这不是因为它们友爱互助,恰好相反,它们需要同类替它们挡子弹:在开阔地躲避捕食者的最好办法就是往同类群里扎;成群出没的习性极大提升了雄性间的性竞争强度,和交配关系中的雌雄比,继而导致雄性发达的第二性征和巨大的性器官。
 开阔草原中,食草动物会成群出没
开阔草原中,食草动物会成群出没
重要的是,特化适应是个不断加速的正反馈过程,策略选择与生理/技能改变轮番相互加强:消化能力越提升,食草策略越受青睐,爪牙越锐利,立体视觉越好,捕猎越有优势,食肉策略越受青睐,反之,草叶在食谱中比例越高,对消化系统的选择压力越强,肉类比例越高,对爪牙和双眼视觉选择压力越强,如此循环,走上一条特化的不归路。
专食与杂食
物种(及更大类元)在特化道路上可能会走得很远,考拉几乎只吃桉树叶,而桉叶以营养低、难消化和毒性强而著称,桉叶精油是强效杀虫杀菌剂,只有考拉和一些负鼠有能力对付;对付桉叶的独特能力让考拉占据了一个极少竞争的生态位,但也失去了很多:考拉代谢率非常低,行动迟缓,反应迟钝,活动范围小,每天睡20个小时,清醒时间几乎全部用来嚼桉叶……幸好澳洲没有擅长爬树的大型食肉动物(比如豹)。
 无尾熊一天能睡20个小时
无尾熊一天能睡20个小时
猫科则走向另一个极端,它们将捕猎禀赋发展到了极致,但由于几乎专吃肉食,其消化系统处理植物的能力严重退化,比如味觉系统丧失了甜味感受器,而后者是辨别植物营养价值的重要手段;无论朝哪个方向,高度特化都降低了物种的适应灵活性,当食物来源随环境条件而改变,或出现新的天敌或竞争者时,很难转向或掉头。
然而并非所有动物都沿食性特化道路走的很远,熊科和猪科都是高度杂食的,犬科和人科的食谱也相当广泛;杂食让这些动物保持了应对环境变化的适应灵活性,所以熊科里才会既有专吃肉食的北极熊,也有吃素——而且几乎只吃极难消化的竹子——的大熊猫,大熊猫从杂食向素食的转变只有两三百万年的历史(和人类转向肉食的时间差不多),这很好的展示了熊科的灵活性。
 吃素的大熊猫
吃素的大熊猫
当然,大熊猫要是在这个特殊生态位下继续进化几百上千万年,或许也会像考拉一样走上高度特化的不归路,反过来说,杂食性可能恰恰体现了这些动物的祖先所走过的进化道路上,环境条件的摆动更频繁,幅度更大,从未提供充足时间让它们完成食性特化。
机会主义者
杂食性代表了一种觅食策略上的机会主义,在素食-肉食这一光谱上,它显然处于中间位置,不过,这个维度对我们理解该策略并没有多大帮助,我们最好从时间分配的角度看待它,即,在面临各种潜在的觅食机会时,将多少时间分配给自己熟悉且擅长处理的食物源,而多少分配给较为陌生的,新颖的,充满未知因素的,价值不明确的食物源。
让我用一个有点类似的生活问题来说明我的意思:我发现自己在超市买食品时经常面临一个两难:日复一日的买同样的食品,难免让人厌倦,感觉自己错过了太多美味,可是尝试新鲜事物的风险也很高,以我个人经验,其中大部分会让我失望,很多最后进了垃圾桶,所以必须作出权衡:将多少预算分配给新食品?多年前我还不会做饭时,在选择餐馆上也面临同样权衡:每十顿饭里几顿留给熟悉的饭馆,几顿用来探索新饭馆?
 要买经常吃的口味?还是尝鲜?
要买经常吃的口味?还是尝鲜?
一旦我们转换到守旧-探索这一维度上,便发现,原来杂食动物才是策略上的激进分子,它们随时准备捕捉任何出现在面前的新机会,而不是一心专注于自己最熟悉擅长的食物源上;所谓机会主义,就是对特定食物源较少持有内在偏好或固有习惯,对新食源总是持开放态度,某一时刻作何选择,全看哪个机会在此时此刻的有着最高预期收益。
这听起来简单,实则对动物的某些禀赋极具挑战,诚然,杂食性对特定捕食技能——诸如鹰的锐眼,猫的柔韧性,鳄鱼的咬合力——没有那么强的选择压力,可是对综合感官和一般认知能力的要求却很高,因为它要求动物在不断面临新情境、新食源的条件下能够良好辨别物体种类与数量,评估其可食性、营养价值和中毒风险,以及面对不同竞争者时的获胜可能性,正因此,猪、熊、狗普遍有着较高的智力,鸟类中的杂食冠军乌鸦也以高智力出名,更别提人科了。
 乌鸦以高智商出名
乌鸦以高智商出名
不妨再以人类职业倾向作类比,许多人偏爱一份稳定职业,有着相对固定的收入,就像食草动物,也有些人是命中注定的连续创业者,朝九晚五这种事情对他们是完全不可接受的,他们是食肉动物,但还有另一些人,他们对职业类型没有任何内在偏好,没有好机会时,也能朝九晚五安心打一份工,可一旦机会出现,比如诱人的跳槽机会,激动人心的创业念头,捞笔外快的良机,则决不会轻易放过,他们是杂食动物——一个并不像其名称所显示的那么中庸的类型。